“Enhän minä voi keskustella kanssasi, koska et ole biologi!”, kommentoi suorasukaiseen tyyliinsä eräs Suomen oloissa huomattava pohjaeläintutkija, kun en vasta-alkaneena vesihyönteisharrastajana tiennyt mikä on sulkahyttynen. Vuosien vieriessä erikoistuin tutkimaan semiakvaattisia ja terrestrisiä hyönteisiä, mutta muutama vuosi sitten palasin ikään kuin juurilleni vesihyönteisten maailmaan. Tässä kirjoituksessa kerron, mitä ovat sulkahyttyset ja mitä uutta olen niiden taksonomiasta, levinneisyydestä ja ekologiasta saanut selville.
Sulkahyttyset ovat keijustohyönteisiä
Kuten heimon nimestä voi päätellä, ovat sulkahyttyset (Chaoboridae) kaksisiipisiä hyönteisiä (Kuva 1). Ne ovat vertaimevien hyttysten (Culicidae) niin sanottu sisarryhmä, eli niillä on lähin yhteinen kantamuoto n. 200 miljoonan vuoden päässä evoluution historiassa. Hyttyset kehittyivät asuttamaan pääasiassa pieniä seisovia vesiä, kuten kausikuivia lammikoita ja sadeveden täyttämiä puiden onkaloita, ja niiden aikuiset naaraat imemään verta selkärankaisista eläimistä. Sulkahyttyset valitsivat toisen tien: niiden toukista tuli petoja ja mestareita elämään suurissakin järvissä. Sulkahyttyset ovatkin ainoita planktonhyönteisiä, eli sellaisia jotka ovat erikoistuneet elämään seisovien vesien keijustossa. Vaikka useita sulkahyttyslajeja on liitu- ja jurakauden fossiiliaineistoissa, ovat useat kehityslinjat kuolleet sukupuuttoon ja muinaisesta runsaudesta on jäljellä vain rippeet. Sulkahyttyslajeja on maailmanlaajuisesti vain 50, siinä missä hyttysiä on yli 3500. Suomesta on toistaiseksi ainoana maana Euraasiassa havaittu kaikki mantereen lajit, joita on yhteensä 11.

Kuva 1. Suomessa esiintyy sulkahyttysiä kolmessa suvussa: Cryophila, Mochlonyx ja Chaoborus. Kuva A. Rinne.
Heimolle yhteisiä tunnusmerkkejä ovat erityisesti toukkien tuntosarvet, jotka ovat muuntuneet liikuteltaviksi pyyntielimiksi, sekä uimarakot, joiden kaasupaineen avulla toukka voi säätää sijaintiaan vesipatsaassa. Kotelon nopean uinnin mahdollistavat perämelat, jotka ovat hyttysten ja sulkahyttysten yhteinen ominaisuus; vastaavaa rakennetta ei ole muilla hyönteisillä. Suomessa esiintyvät sulkahyttyset kuuluvat kolmeen eri sukuun, Cryophila, Mochlonyx ja Chaoborus. Cryophila ja Mochlonyx ovat erikoistuneet elämään pienissä lammissa ja kausikuivissa lammikoissa ja ne ovat tärkeitä hyttystoukkien saalistajia. Kaikki lajit ovat munatalvehtijoita ja niiden toukat esiintyvät tyypillisesti suhteellisen lyhyen aikaa keväällä ja alkukesällä. Molempien sukujen toukilla on myös hengitysputket, joilla ne voivat ottaa ilmakehän happea. Chaoborus-toukat sen sijaan saavat tarvitsemansa hapen suoraan vedestä ihon läpi. Ne ovat myös läpinäkyviä, siinä missä Cryophila ja Mocholonyx ovat oranssinruskeita tai hieman läpikuultavia. Chaoborus-lajit elävät joko järvissä tai lammissa, ja niiden toukat talvehtivat; ainoa poikkeus on munana talvehtiva pohjoinen erikoisuus Ch. nyblaei. Toukkien läpikuultavuudesta tulee myös heimon kansanomainen nimi phantom midges, eli kummitushyttyset; suomenkielinen nimi ehkä viittaa kotelon sulkamaisiin perämeloihin.
Alhaista lajimäärää kuitenkin kompensoi sulkahyttysten suuret paikalliset runsaudet. Eräitä järvilajeja voidaan parhaimmillaan tavata jopa kymmeniä tuhansia toukkia neliömetriä kohti, joten yksittäisten järvien populaatiot lasketaan helposti sadoissa miljoonissa yksilöissä. Koska toukat ovat melko suurikokoisia (pituus täysikokoisena 7-22 mm) petoja, on niillä myös suuri vaikutus keijuston koostumukseen ja täten ravintoverkkoon. Suomessa on vain yksi järvilaji (Chaoborus flavicans), muut sulkahyttyset elävät toukkina erilaisissa kalattomissa lammissa; kalojen saalistus onkin tärkein lajien vesistökohtaista esiintymistä selittävä tekijä. Sulkahyttysiä pidetään yleisesti ottaen ihmiselle hyödyllisinä, koska ne saalistavat hyttystoukkia ja ovat mm. kalojen ravintokohteita. Toisaalta Chaoborus-toukat voivat joskus olla haitallisia kalanpoikasten kasvatuslammissa, koska ne saattavat syödä pienpoikasia! Kaliforniassa Chaoborus astictopus on esiintynyt eräillä järvillä niin runsaana, että sen suuret aikuisten massaesiintymät ovat häirinneet rannoilla asuvia ihmisiä ja tätä kautta alentaneet kiinteistöjen arvoa. Tästä syystä tätä lajia on paikallisesti yritetty hävittää larvicideilla eli toukkia tappavia myrkkyjä käyttämällä.
Järvissä esiintyvillä sulkahyttysillä on eräitä sopeumia, joiden avulla ne kestävät kalojen saalistusta. Ensinnäkin, toukat voivat aistia kalojen läsnäolon kemiallisesti. Kokeellisesti on osoitettu, että kalattomassa vedessä eläneet C. flavicans toukat laskeutuivat välittömästi pohjalle ja kaivautuivat sedimenttiin, mikäli niiden altaaseen lisättiin vettä kala-altaasta. Järvilajien toukat ovat päivisin joko kaivautuneita pehmeään pohjasedimenttiin tai ne ovat hapettomassa alusvedessä; auringon laskettua ne nousevat päällysveteen syömään pieniä vesikirppuja ja hankajalkaisia äyriäisiä. Tällä käytöksellä ne voivat varsin hyvin välttää näköaistin avulla saalistavien kalojen ahnaat suut. On melko huomionarvoista, että reilun senttimetrin pituiset toukat voivat vuorokauden aikana nousta useiden kymmenien metrien matkan vesipatsaassa ja laskeutua jälleen takaisin pohjan kylmyyteen. Afrikan suurissa järvissä toukkien vertikaalinen vaellus voi olla jopa 200 metriä suuntaansa. Vaellus vesipatsaassa on mahdollista hengityselimistöstä erilaistuneiden uimarakkojen avulla, jotka ovat parilliset rakenteet keskiruumiissa ja takaruumiin peräpäässä. Uimarakkojen pinnalla on pigmenttisoluja (kromatoforisia läiskiä), joiden ansiosta Chaoborus-toukkamassat voi havaita veneen kaikuluotaimesta. Lampilajien toukkien uimarakot eivät kestä suuria paineenvaihteluita ja tästä syystä ne eivät pysty tekemään vertikaalisia vaelluksia. Koska lampilajit eivät voi paeta kaloja, ne välttävät niitä. Naaraat pystyvät aistimaan veden pinnalla kemiallisten signaalien, ns. kairomonien avulla, onko vedessä kaloja. Näin ollen naaraat voivat valikoivasti munia kalattomiin vesiin.
Suomen sulkahyttysiä tutkimassa
Ennen vuotta 2013 olin pitänyt sulkahyttysiä erittäin hyvin tunnettuina, siis sellaisena ryhmänä josta tuskin voisi löytää mitään erityisen uutta tai mielenkiintoista. Vuonna 2013 olin erään päivänkorentolajin kartoittamisen yhteydessä Savukoskella ottanut talteen kalattomasta lammesta suurikokoisia sulkahyttystoukkia, ihan vain päähänpistosta. Koska en tuolloin tuntenut lajistoa, lähetin toukat Lauri Paasivirralle määritettäväksi. Suomen kokeneimmalta pohjaeläintutkijalta tulikin pian vastaus, että näytteissä oli ollut Ch. nyblaei, joka oli kerätty Suomesta aikaisemmin vain Utsjoen Nuorgamista, 1800-luvun lopulla ja vuonna 1960. Koska kyseessä oli näyttävä, yli 20 mm pitkä suurikokoinen laji (Kuva 2), ajattelin sen myös olevan suuri harvinaisuus: ehkä laji esiintyy vain Savukoskella Maanselän vedenjakajan itäpuolella sekä aivan pohjoisimmassa Lapissa. Tästä havainnosta innostuneena keräilin vuonna 2014 muutamilta paikoilta Savukoskelta toukkia ja aloitin perehtymisen niiden määrittämiseen. Mitenkään systemaattisesti en sulkahyttysiä tuossa vaiheessa tutkinut, vaan sivubisneksenä muiden töiden ohella. Olin kuitenkin ottanut keräämistäni näytteistä kudosnäytteitä ja toimittanut niitä FINBoL-hankkeen (Finnish Barcode of Life) puitteissa kansainväliseen BOLD-lajitunnistetietokantaan (Barcode of Life Database). Tästä tietokannasta ja siinä olevista havainnoista tulisi jatkossa olemaan minulle suuri hyöty.

Kuva 2. Chaoborus nyblaei on pohjoinen harvinaisuus, joka tunnetaan vain Norjasta, Ruotsista ja Suomesta. Laji on Euroopan suurikokoisin ja se asuttaa kausikuivia ja pysyviä kalattomia lampia. Kuva A. Rinne.
Vuonna 2017 keräilin sulkahyttysiä hieman laajemmin, esimerkiksi Rovaniemen seudulta ja Tuntsan erämaasta. Tuona kesänä sain saaliiksi ensimmäisen kerran mm. Cryophila lapponica-lajin, jota pidin vielä tuolloin hyvinkin harvinaisena. Kuitenkin vasta 2018 aloin kunnolla selvittämään lajiston esiintymistä Lapissa ja eteläisessäkin Suomessa. Otin tavoitteekseni tehdä yhteenvedon Suomen lajistosta sekä tehdä runsaasti kuvitetun määrityskaavan toukkiin, koteloihin ja aikuisiin. Tuona maastokautena retkeilin Tervolassa, Rovaniemellä, Posiolla, Kittilässä, Savukoskella, Sallassa, Inarissa ja Utsjoella. Lisäksi pyysin muutamalta kaverilta näytteitä, ja sainkin niitä esimerkiksi Salosta, Espoosta, Urjalasta, Tampereelta, Petäjävedeltä, Kiteeltä ja Saariselältä. Kemijoen itäpuolella, lähellä Rovaniemen ja Tervolan rajaa, selvitin muutaman lammen lajistoa alku- ja loppukesän välisenä aikana. Näillä lammilla käytin mm. kuoriutumispyydyksiä (Kuva 3), jotka ovat kelluvia, kartiomaisia lauttoja. Pyydyspurkki sijaitsee kartion huipussa ja sinne päätyvät kaikki pyydyksen kohdalla kuoriutuneet hyönteiset, kuten hyttyset, surviaissääsket, vesiperhoset ja niukalti polttiaiset. Lisäksi keräsin toukkia ja kasvatin niitä myös aikuisiksi. Suuri apu minulla oli Erkka Laineesta, joka teki Itä-Lapin aineistosta pro gradu-tutkielmansa Jyväskylän yliopistoon.

Kuva 3. Kuoriutumispyydys vuonna 2018 Rovaniemen ja Tervolan rajaseudulla. Kuva J. Salmela.
Vuosi 2018 oli tuloksellisesti menestyksekäs. Keräsin aineistoa lopulta yli 90 lammelta ja tietämys useiden lajien levinneisyydestä parani merkittävästi. Rariteettina pitämäni Ch. nyblaei osoittautui suhteellisen tavalliseksi Itä-Lapin kalattomissa lammissa, joista osa oli kausikuivia. Lisäksi Ch. nyblaei esiintyi melko tavallisena Rovaniemen ja Tervolan rajoilla olevalla ylänköalueella, eli sen tunnettu levinneisyysalue laajeni huomattavasti etelään. Niin ikään Cr. lapponica osoittautui suhteellisen tavalliseksi pienten lampien ja lammikoiden lajiksi. Lajien paljastuminen yleisiksi on siinä mielessä kummallisia, että molemmat ovat varsin suurikokoisia ja näyttäviä, ja ne on helppo määrittää lajitasolle. Selityksiä näennäiselle harvinaisuudelle ovat kalattomien lampien hyönteisten vähäinen tutkimus ja lajien pohjoinen levinneisyys.
Tuon kesän hienoimpia tuloksia oli kuitenkin Suomelle uuden Mochlonyx-lajin identiteetin varmistuminen. Tämä laji oli ollut mysteeri, koska se ei DNA-lajitunnisteprofiililtaan sopinut kumpaankaan Suomesta (tai Pohjoismaista) aiemmin tunnettuun lajiin. Euroopasta kylläkin tunnettiin kolmaskin laji, vasta vuonna 1982 Hollannista kotelon ja aikuisen perusteella kuvattu M. triangularis, jota ei oltu tavattu muualta sen koommin. Olisi kuitenkin ollut mahdollista, että pohjoinen mysteerini olisi ollut jokin pohjois-amerikkalainen laji tai jopa uusi, joten asia ei voinut ratketa kuin saamalla aikuisia yksilöitä tutkittavaksi. Minullahan ei tätä kesää ennen ollut kuin toukkia kerättynä. Mutta miten saada aikuisia? Se onnistuu helpoiten kasvattamalla (Tietolaatikko).
Tietolaatikko. Sulkahyttysten kasvattaminen on verrattain helppoa. Maastosta kerätty täysikasvuinen ns. IV-vaiheen toukka tai kotelo (pupa) tuodaan sisätiloihin ja asetetaan omaan kasvatusastiaan. Astiaksi sopii parhaiten viritelmä, jossa on kaksi muovista näytepurkkia, tilavuudeltaan noin 2 dl, toisen toimiessa vesialtaana ja toisen kattona. Näiden välissä on purkin kansi, johon on tehty halkaisijaltaan reilun kahden cm reikä. Kasvatuksessa olevalle toukalla voi antaa ravinnoksi pieniä äyriäisiä (esim. vesikirppuja) tai hyttystoukkia. Yksilön koteloituessa se pudottaa vanhan toukkanahkan pääkapseleineen pois päältään. Toukkanahka otetaan talteen etanoliin pieneen näyteputkeen ja tässä vaiheessa annetaan kasvatukselle oma yksilöllinen koodi, jolla pystytään yhdistämään saman yksilön eri kehitysvaiheet; tämä on tarpeen varsinkin jos kasvatettavia yksilöitä on useita kymmeniä. Kotelovaihe kestää huoneenlämmössä muutaman päivän. Aikuinen yksilö kuoriutuu veden pinnalla ja kotelonahka jää pinnalle kellumaan. Tämä on kasvatuksen kriittisin vaihe: mikäli kasvattaja ei ole tarkkana, pääsee siivellinen aikuinen helposti karkuun. Tästä syystä aikuinen on varmin kerätä, kun se lepää kattona olevassa purkissa tai kannen yläpuolella. Tällöin on helppo ujuttaa paperi katon alta väliaikaiseksi lattiaksi, jolloin katto ja sen sisällä oleva sulkahyttynen voidaan nostaa pois altaan päältä. Tässä vaiheessa etsitään kattoon sopiva ehjä kansi ja se asetetaan paperin päälle purkin suulle. Kun paperi vedetään välistä varovasti pois, jää aikuinen satavarmasti vangiksi purkkiin. Kotelonahkan voi säilöä samaan putkiloon toukkanahkan kanssa ja aikuiselle annetaan sama näytetunnus kuin toukkanahkalle. Aikuinen viedään hetkeksi pakkaseen ja tämän jälkeen joko neulataan tai säilötään samaan putkiloon nahkojen kanssa.
Melko pian havaitsin, että samoilla lammilla ja lammikoissa esiintyy kahta Mochlonyx-lajia. Fenologisesti selvästi aikaisempi M. velutinus oli näistä kahdesta suurempi ja aikaisemmin Suomesta tunnettu. Esimerkiksi Rovaniemellä tämä laji kuoriutui jo kesäkuun alkupuolella. Myöhäisempi laji oli toukkana hieman pienempi ja vaaleampi, aikuisten lentoajan ollessa vasta heinäkuussa. Saatuani ensimmäiset koiraat tästä myöhäisemmästä pystyin nopeasti määrittämään niiden kuuluvan lajiin M. triangularis. Ajattelin tämän olevan ensimmäinen ja viimeinen Suomelle uusi laji tästä heimosta. Ilokseni sain huomata olleeni väärässä.
Chaoborus flavicans: kuinka yhdestä lajista tuli kolme
Vuoden 2018 maastokauden kääntyessä syksyksi aloin määrittämään sesongin aikana kertynyttä laajaa aineistoa. Erkan graduaineistossa Savukosken Törmäojan lähettyviltä oli yksi lampi, jossa oli runsaasti Ch. flavicans-tyyppistä toukkaa. Kuten edellä totesin, on Ch. flavicans sulkahyttystemme ainoa järvilaji, mutta se voi myös joskus esiintyä kalattomissa, pienemmissä lammissa. Kuitenkaan Lapissa se ei juuri koskaan esiinny pienvesissä ja ehkäpä tästä syystä päätin katsoa törmäojalaisia hiukan tarkemmin. Chaoborus flavicans on helppo tuntea keskenkasvuisenkin toukan perusteella, koska sillä ainoana sukunsa lajina mandibelin (eräs suuosa) ns. pikkuhammas eli III hammas sijaitsee täsmälleen II ja IV hampaan välissä (Kuva 4). Tarkastelussa olevilla toukilla tämä pikkuhammas oli flavicans-tyyppiä, mutta mandibelin ulommalla reunalla olevat sivuhampaat näyttivät varsin jykeville. Jostain alitajunnasta mieleeni tulvahti, että olen nähnyt kuvan tästä hammastyyppistä jossakin julkaisussa. Pienen pdf-tiedostojen penkomisen jälkeen löysin etsimäni, eli professori Derek J. Taylorin tutkimusryhmän artikkelin vuodelta 2008. Taylor oli Alaskasta havainnut toukkatyyppiä, joka erosi perus-flavicans-lajista juuri sivuhampaiden koon perusteella. DNA-analyysien mukaan alaskalaista cf. (=confer, vertaa) flavicans-lajia esiintyi myös muualla Pohjois-Amerikassa ja Japanissa. Lisäksi cf. flavicans erosi huomattavan paljon DNAltaan lajista C. flavicans, määrällä joka on tyypillistä jo useita miljoonia vuosia sitten tapahtuneelle lajiutumiselle.
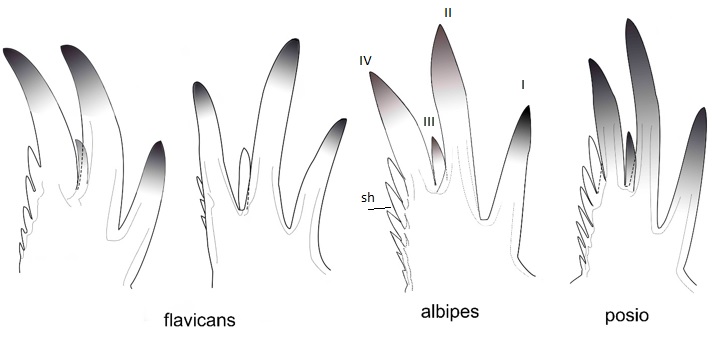
Kuva 4. Chaoborus flavicans-lajiryhmän lajien mandibelit. sh=sivuhampaat, I-IV=hampaiden numerointi. Kuva J. Salmela.
Samankaltaista cf. flavicans toukkatyyppiä määritin myös Posion Peuralamminharjulta sekä Utsjoelta Tenojoen laaksosta vuoden 2018 keräyksistä. Otin toukista kudosnäytteitä ja sain FINBoLin kautta niiden DNA-lajitunnisteet 2019 alkuvuodesta. Yllätykseni oli suuri, kun flavicans-tyyppiset “oksat” eli yksilöt sijoittuivat “määrityspuussa” kolmeen selvästi erottuvaan haaraan: minulla oli todennäköisesti Suomesta ei vain kaksi vaan jopa kolme lajia! Chaoborus flavicans ja kaksi muuta, sillä cf. flavicans jakaantuikin kahteen lajiin. Tässä vaiheessa saatoin vain todeta, että Suomen lajiston revisio ei ollutkaan niin helppo homma kuin olin ajatellut.
Keväällä 2019 päätin, että havaittu taksonominen sekamelska on selvitettävä kunnolla. Selvitettäviä asioita oli useita: mikä on alaskalainen cf. flavicans ja onko se samaa lajia kuin toinen minun lajeistani (joista aloin käyttämään työnimiä “posio” ja “törmäoja” löytöpaikkojen mukaan); onko näillä lajeilla jo ns. saatavilla olevat nimet flavicans-lajin synonyymien joukossa vai ovatko ne uusia; mikä on näiden taksonien levinneisyys ja ekologia Suomessa? Ensimmäiseksi minun pitäisi saada kasvatettua toukista aikuisia, eli minun olisi kesällä 2019 käytävä edellisen vuoden lammilla hakemassa eläviä toukkia. Aikuisten välisten, varsinkin koiraiden sukupuolielinten, rakenne-erojen perusteella voisin varmistua, että DNAn erottamat lajit ovat ns. oikeita lajeja, eikä kyse olisi vain toukkien lajinsisäisestä vaihtelusta. Tämän jälkeen minun pitäisi käydä läpi vanhaa taksonomista nimistöä ja nimiin liittyviä tyyppiyksilöitä. Chaoborus flavicans-lajilla oli neljä nuorempaa synonyymiä, eli myöhemmin annettua nimeä jotka tarkoittavat samaa lajia. Näiden nimien tyyppiyksilöistä kolme löytyisi museoista Yhdysvalloista, mutta yhden Euroopasta annetun nimen tyyppiyksilöiden sijaintia ei tunnettu. Vanhojen keräyspaikkojen ohella yritin etsiä näitä lajeja myös uusilta lammilta.
Aloitin maastokauden toukokuun alussa retkellä, jonka aikana keräsin aineistoa mm. Kouvolasta, Rautjärveltä, Savolinnasta, Joroisilta, Vieremältä ja Muhokselta. Myöhemmin toukokuussa kävin myös Suomussalmen Hossassa, jonka lampiluonto on suorastaan häkellyttävän monimuotoista. Kasvatukset sekä kesän maastotyöt onnistuivat erittäin hyvin, ja pian hallussani olikin useiden kymmenien yksilöiden “täydelliset” sarjat, joissa oli saman yksilön toukka- ja kotelonahka sekä aikuinen. DNAn ennustamat lajit erosivat myös aikuisten morfologian perusteella, joten olo oli aika juhlava kun olin saanut varmistettua kahden Suomelle, ja ehkä jopa tieteelle, uuden lajin olemassaolon. Chaoborus “törmäojan” tunnettu levinneisyysalue laajeni huomattavasti, sillä sitä tuli mm. Kouvolasta ja Kittilästä. Chaoborus “posio” sen sijaan näyttäisi olevan pohjoinen laji, sillä sen eteläisimmän populaation havaitsin Hossan kansallispuistosta.
Vuoden 2019 syksyllä aloin tekemään toukkien, koteloiden ja aikuisten mikroskooppisia mittauksia, joita tarvittaisiin tieteellisesti pätevän artikkelin tarpeiksi. Lisäksi olin yhteydessä useisiin ulkomaisiin museoihin ja tutkimuslaitoksiin, joista pyysin edellä mainittujen tyyppiyksilöiden ohella sulkahyttysiä lainaksi tutkimustani varten. Onneksi sain haluamani lainat joulu-helmikuun aikana, ennen kuin koronakriisi sulki museot. Melko nopeasti varmistui, että “posio” oli todellakin uusi laji, mutta “törmäojan” identiteetin selvittäminen vaati kaikkien kivien kääntämistä. Tutkimieni tyyppiyksilöiden joukossa oli vuonna 1903 kuvattu Corethra albipes, jonka holotyyppinaarasta en varsinaisesti tutkinut lainkaan. Siitä oli nimittäin enää jäljellä vain yksi lasille preparoitu siipi, ja minulle riitti että sain kuvan siitä nähdäkseni. Sayomyia rotundifolia-lajin holotyyppiä ei oltu nimetty vuoden 1904 alkuperäisessä kuvauksessaan, joten kyseessä oli ns. syntyyppisarja, joka koostui neljästä lasipreparaatista. Näistä yhdessä oli koiraan takaruumin kärki, ja näin ollen myös määrityksen kannalta tärkeät sukupuolielimet. Kolmas tyyppiyksilö oli vuonna 1924 kuvatun Ch. eluthera-lajin lektotyyppi, joka niin ikään oli lasille preparoitu koiraan takaruumis. Neljännen synonyymin (Ch. alpinus, vuodelta 1938) tyyppiyksilöitä kyselin eri museoista Saksasta, mutta niiden sijaintia ei kukaan tunnu tietävän.
Chaoborus eluthera-nimen oli helppo vahvistaa olevan edelleen Ch. flavicans-lajin synonyymi, samoin Ch. alpinus, kiitos alkuperäisen kuvauksen hyvien kuvien. Sitten huomasin, että S. rotundifolia on selvästi sama laji kuin “törmäoja”. Ongelma oli Co. albipes, koska sen tyyppiyksilöstä ei ollut enää mitään hyötyä. Luin kuitenkin tarkasti lajin alkuperäisen kuvauksen. Sen mukaan takaruumiin jaokkeiden sivuilla pitäisi olla tummia läiskiä (“specks”). Nämä läiskät ovat todellakin tyypillisiä pohjois-amerikkalaisille “törmäoja” yksilöille (Kuva 5), joten suomalainen “törmäoja” on sama kuin Atlantin takainen albipes. Tällä lajilla on laaja levinneisyys ympäri pohjoisen pallonpuoliskon, sillä olen nyt voinut varmentaa sen esiintyvän Norjassa, Suomessa, Venäjällä, Japanissa, Yhdysvalloissa ja Kanadassa. Näin ollen Chaoborus flavicans lajikompleksi koostuu vähintään kolmesta lajista, joista kaksi on levinneisyydeltään holarktisia (flavicans, albipes) ja yksi euraasialainen (toistaiseksi vain Suomesta tunnettu “posio”).

Kuva 5. Chaoborus flavicans (2 vasemman puoleista) ja Ch. albipes (keskimmäinen ja kaksi oikealta) aikuisten takaruumiit päältä katsottuna. Lajilla flavicans on tummat juovat tergiiteissä 2-5, mutta lajilla albipes läiskät sivuilla ja keskellä, ei yhtenäistä juovaa. Pohjoisesta (Pohjoismaat, Alaska, Hokkaido) kerätyillä yksilöillä, kuten oikealla, takaruumiin väritys on tumma ja läiskät erottuvat heikommin. Kuva J. Salmela.
Kalattomien lampien tuntematon monimuotoisuus
Chaoborus flavicans on laajalle levinnyt, runsas laji, jonka ekologia ja morfologia tunnetaan poikkeuksellisen hyvin. Miten on mahdollista, että kaksi lähilajia havaitaan tai kunnollisesti tunnistetaan vasta nyt? Uskoisin kahden syyn nousevan selittäviksi tekijöiksi. Ensinnäkin Ch. flavicans on selvästi järvilaji, ja sillä on todennäköisesti lajiryhmänsä ainoana kehittynyt kyky tehdä vertikaalisia vaelluksia vesipatsaassa. Kaksi muuta lajia elävät kalattomissa pienvesissä, joita on tutkittu vähemmän. Toinen tekijä on aikuisten yksilöiden lyhyt elinikä ja suhteellisen vaikea saatavuus. Sulkahyttysiä on määritetty enimmäkseen toukkina, ei aikuisina. Pitäisin todennäköisenä, että joku muukin on minua ennen Euraasiasta kerännyt kalattomista lammista toukkia ja määrittänyt niiden olevan lajia Ch. flavicans, mutta ovat oikeasti kuuluneet lajiin Ch. albipes. Ehkä kukaan ei ole tullut huomanneeksi mandibelin isojen sivuhampaiden kokovaihtelua tai tajunnut sen taksonomista merkitystä; on ehkä vain oletettu, että tämä kuuluu normaalin vaihtelun piiriin.
Havaintoaineiston valossa näyttäisi sille, että Ch. albipes on tavallinen laji. Esimerkiksi keväällä 2020 vanha opiskelutoverini ja luontoalan moniosaaja Oskari Härmä löysi useita aikaisemmin tuntemattomia populaatioita Lounais-Hämeestä. Ei sen pohjoinen lähilaji “posiokaan” mikään suuri harvinaisuus ole, mutta lajien biologiassa on eroavaisuuksia. Siinä missä Ch. albipes elää pienissäkin lammissa, olen tavannut “posiota” vain hieman isommista lammista (pinta-ala vähintään 0,25 ha). Epäilen, että lajien väliset vuorovaikutukset vaikuttavat siihen, millaisissa vesissä ne voivat elää. Suomessa kalattomien lampien tavallisin sulkahyttynen on Ch. obscuripes, joka on suurempi kuin yksikään flavicans-ryhmän lajeista. Jostain toistaiseksi tuntemattomasta syystä Ch. albipes pärjää sen kanssa pienissä vesissä, mutta “posio” ehkä tarvitsee syvyyttä tai suuremman avovesialueen, jotta se pääsee välttelemään kilpailullisesti vahvempaa lajitoveria.
Vaikka olenkin saanut selville paljon uutta tietoa Suomen lajistosta, olisi lajien levinneisyyksien ja elintapojen selvittämisessä vielä paljon tehtävää. Esimerkiksi laajalle levinnyt M. fuliginosus näyttäisi puuttuvan isompien lampien yhteisöistä ja elävän todella pienissä vesissä, kuten lähteissä ja suorimmissä. Tarkkaan ei tiedetä, millainen on sen elinkierto, esimerkiksi voisiko lajilla olla kaksi sukupolvea vuodessa. Cryophila lapponica-lajia on noin 90 vuotta sitten tavattu läheltä Pietaria, eli varsin etelästä verrattuna Suomen eteläisimpiin havaintopaikkoihin Muhoksella ja Kuusamossa. Pitäisin mahdollisena, että lajin levinneisyys ulottuu idässä ainakin Pohjois-Karjalaan ja Kainuuseen. Laji olisi myös erinomainen seurannan kannalta, koska pienetkin toukat pystyy määrittämään maastossa silmämääräisesti. Mochlonyx triangularis oli alkuun tiedossani vain parilta paikalta Savukoskelta, mutta laji on osoittautunut Lapissa erittäin yleiseksi ja sitä esiintyy myös eteläisessä Suomessa; lyön tuopillisen konjakkia vetoa, että laji esiintyy lähes koko Fennoskandian alueella. Chaoborus nyblaei on ainoa munana talvehtiva laji suvussaan; ei kuitenkaan tiedetä, muniiko laji luonnossa veteen vai maalle. Lisäksi se melko suurella varmuudella esiintyy etelämpänä (Koillismaa, Kainuu) kuin mitä tällä hetkellä tiedetään, mutta pohjoisena lajina se voi taantua ilmastonmuutoksen seurauksena. Toiveissani onkin, että Suomen lajiston revision ja määrityskaavojen julkaisun jälkeen hyönteisharrastajilla herää kiinnostus havainnoida sulkahyttysiä ja samalla parantaa tietämystä niiden esiintymisestä. Sulkahyttyset ovat omiaan vaikkapa kouluihin ympäristöopin tunneille, koska ne eivät esim. sammakoiden tapaan ole rauhoitettuja ja niitä voi helposti tarkkailla pienissä vesiastioissa.
Kirjallisuus:
Berendonk TU 1999 Influence of fish kairomones on the ovipositing behavior of Chaoborus imagines. Limnology and Oceanography 44 (2), 454–458.
Berg, K. 1937. Contributions to the biology of Coretha Meigen (Chaoborus Lichtenstein). Biologiske Meddelser 13: 1-101
Borkent, A. (1979) Systematics and bionomics of the species of the subgenus Schadonophasma Dyar & Shannon (Chaoborus, Chaoboridae, Diptera). Quaestiones Entomologicae, 15, 122–255.
Borkent A 2014 World catalog of extant and fossil Chaoboridae (Diptera). Zootaxa 3796 (3): 469–493.
Cook, E.F. (1956) The Nearctic Chaoborinae (Diptera: Culicidae). University of Minnesota Agricultural Experiment Station Technical Bulletin, 218, 1–102.
Dawidowicz P, Pijanowska J & Ciechomski K (1990) Vertical migration of Chaoborus larvae is induced by the presence of fish. Limnology and Oceanography 35: 1631-1637.
Dupuis D, Svensson J-E & Taylor DJ (2008) The cryptic origins of environment-indicating phantom midges (Chaoborus). Limnology and Oceanography 53: 236–243.
Garcia, E., & Mittelbach, G. (2008). Regional coexistence and local dominance in Chaoborus: species sorting along a predation gradient. Ecology, 89(6), 1703-1713.
Gosselin A & Hare L 2003 Burrowing behavior of Chaoborus flavicans larvae and its ecological significance. Journal of the North American Benthological Society 22: 575–581.
Hirvenoja M (1961) Weitere Studien über Chaoborinen (Dipt., Culicidae). Beschreibung der Larve und der Puppe von Chaoborus (Schadonophasma) nyblaei. Zett. - Annales Entomologici Fennici 27:77-83.
Ogawa, J.R. (2007) Phylogeny of the “Chaoboriform” genera. Doctor of Philosophy thesis, Oregon State University, Oregon, xiv + 256 pp.
Parma, S. (1971a) Chaoborus flavicans (Meigen) (Diptera: Chaoboridae): an autecological study. Doctor of Philosophy thesis, University of Groningen, Groningen, 128 pp.
Saether OA (1967) Variation within immature stages of Chaoborus flavicans (MEIG.) (syn. Chaoborus alpinus Peus syn. nov.). Internationale Revue der gesamten Hydrobiologie 52: 573-587.
Saether, O.A. (1972) VI. Chaoboridae. In: Zooplankton der Binnengewässer. 1. Tiel. Binnengewässer, 26, pp. 257–280.
Sikorowa, A. (1973) Morfologia, biologia i ekologia gatunkow rodzaju Chaoborus Lichtenstein (Diptera, Chaoboridae) wystepujacych w Polsce. Zeszyty Naukowe Akademii Rolniczo-Technicznej w Olsztynie, 105, 1–121.
Taylor DJ, Ballinger MJ, Medeiros AS & Kotov AA (2016) Climate‐associated tundra thaw pond formation and range expansion of boreal zooplankton predators. Ecography 39: 43-53.
Valle KJ 1930 Über das Auftreten von Mysis relicta und Corethra plumicornis während des Sommers in einigen Seen von Südost-Finnland. Archiv für Hydrobiologie 21: 483-492.
Ei kommentteja:
Lähetä kommentti